Because the climate is changing quickly and trees tend to be genetically adapted to past climate conditions, managers across the globe have begun to implement climate-adapted seed transfer, wherein tree seedlings are planted in locations where current and anticipated climate conditions better match their genetic adaptations. This typically involves planting seedlings at higher elevations or more poleward latitudes than their seed source, in order to track climate change.
Table 1. Some sources of uncertainty inherent to seed selection and climate adaptation.
| Type | Description |
|---|---|
| Geographic | Climate uncertainty arising from use of imprecise and climatically heterogenous spatial units (e.g., seed zones). Geographic uncertainty in both planting and seed-source location can be eliminated by using precise spatial information. However, much of the existing seed inventory in state and federal seedbanks suffers from substantial geographic uncertainty. |
| Future Climate | Uncertainty in what the future climate will be. Two components include (a) climate sensitivity–how sensitive the climate is to GHG-forcing, and (b) emissions pathway–the quantity and timing of future GHG emissions. |
| Model | Uncertainty arising from the inability of statistical models to predict 100% of the variation in outcomes. For instance, climate transfer functions perform well predicting the average outcome of multiple seed sources planted into a new location, but substantial uncertainty remains in the performance of individual seed sources. |
Planting trees necessarily involves uncertainty in survival and growth rates, which in turn means uncertainty in carbon sequestration, timber production, ecosystem services, and habitat value (Table 1). Landowners and managers tend to strongly prefer more reliable reforestation outcomes. In myriad domains, from finance to ecosystem management, diversified portfolio or “bet-hedging” strategies are used to optimize tradeoffs between expected returns and uncertainty in those returns. How can diversified portfolios of seed sources be used to minimize the uncertainty of reforestation outcomes while maximizing forest health and productivity?

Figure 1. Large decreases in the uncertainty of reforestation outcomes may be achieved by planting optimal portfolios of multiple seed sources at reforestation sites. This conceptual diagram, representing results from six million simulations, is based on data from a large seed transfer experiment (O’Neill & Nigh 2011) and estimates of future climate uncertainty derived from global circulation models (Pierce et al. 2014). Left Panel: Selection of optimal portfolios (labeled A-E) relative to the efficient frontier (solid curve). Right Panel: Curves depict probability distributions. Boxes depict medians and interquartile ranges.
To take a first look at this problem I used data from the Illingworth Trial, which planted over 70,000 Pinus contorta seedlings from 140 seed-source locations in 62 common gardens and measured them for 32 years (O’Neill & Nigh 2011). I fit a univariate, asymmetric, gaussian climate transfer function to predict relative tree volume and its uncertainty as a function of mean annual temperature (MAT) transfer distance (site climate – source climate). I considered 26 candidate seed sources–each adapted for optimal performance at a different MAT, and randomly generated 6,026 candidate portfolios each composed of different proportions of each seed source. Based on the results of global circulation models (GCMs) I assumed that the probability of realized MAT at our planting site follows a normal distribution with a standard deviation of 0.8 °C (Pierce et al. 2014). I then ran 1,000 simulations per portfolio to assess the probability distribution of outcomes associated with each portfolio. Based on the outcome of these simulations I selected optimal candidate portfolios that fell near the efficient frontier (Dragicevic et al. 2016). This simplified approach serves as a proof of concept and provides a glimpse of the improved reliability that could be achieved by using diversified portfolios of seed sources in reforestation (Fig. 3).
It appears that the risks associated with the common strategy of planting a single seed source at a reforestation site can be mitigated substantially. Diversified portfolios necessarily produce more reliable outcomes when the performances of the individual components of a portfolio of assets (whether they are trees or financial instruments such as stocks) are less correlated and the number of assets in the portfolio is larger (Markowitz 1952). To illustrate this, consider future climate uncertainty at a given reforestation site. If we plant a single seed source–genetically adapted to the mean expected climate at the site–it will be subject to reduced growth and survival when realized climate conditions vary from our expectation. In contrast, if we instead plant two seed sources, adapted respectively to conditions slightly warmer and cooler than the mean expected climate such that their performance tends to be negatively correlated, the overall outcome will become more reliable. Data from seed transfer experiments indicate that the performance of different combinations of seed sources (in multidimensional climate space) ranges from negatively correlated, to uncorrelated, to positively correlated. Using data from seed transfer experiments, we can construct optimal portfolios of seed sources that maximize both average reforestation outcomes and their reliability (Weng et al. 2013).
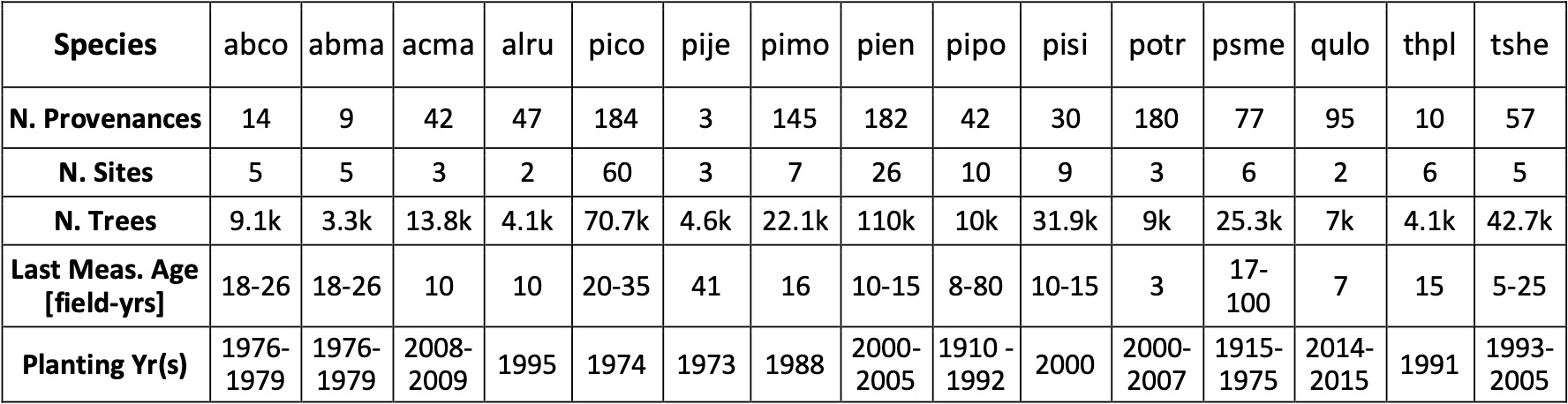
Table 2. Characteristics of extant common garden (AKA “provenance test” or “seed transfer experiment”) datasets that may be suitable to use in this component of the project. 4-letter species codes are based on genus and species names. For example, abco = Abies concolor, pico = Pinus contorta.
While the analysis outlined above makes a few simplifying assumptions, we plan to expand on this approach to more fully and accurately predict the probability distribution of outcomes associated with optimally diversified portfolios of seed sources. This work is funded by the University of California Office of the President. Whereas the simplified approach accounts for only one dimension of climate, the full approach will account for seed adaptations to, and uncertainty in, multiple dimensions of climate (e.g., seasonal variation in temperature, precipitation, etc.). Whereas the simplified approach does not account for how uncertainty varies as a function of the number of trees planted, the full approach will leverage available data to account for this relationship. Whereas the simplified approach makes assumptions about the shape and collinearity of probability distributions, the full approach will better account for these important factors. Whereas the simplified approach does not account for geographic uncertainty in the origin of seeds (e.g., seed zones), the full approach will incorporate this uncertainty. We plan to leverage much of the available data from extant common garden datasets planted in western North America in these analyses (Table 2). Finally, we will seek to develop efficient algorithms that can identify and propose optimal portfolios of available seed sources in real-time (i.e., in response to user inputs to an online tool such as the Climate-Adapted Seed Tool).
In the meantime, given apparently ubiquitous preferences for reliable outcomes (Dragicevic et al. 2016), we suggest that reforestation projects use at least a few different seed sources per species planted. Of course, each of these seed sources should also be selected to match the genetic adaptations of the seed source with the climate of the planting site. For example, with the current version of CAST, this could be accomplished by using the top three candidate seed sources for the climate conditions of your planting site. Alternatively, a bit more diversification could be introduced by varying your expected climate conditions. For instance, instead of selecting seeds adapted to the expected climate ~20 years in the future at your site, you could select one seed lot adapted to ~20 years into the future, another adapted to expected conditions ~15 years in the future, and another adapted to expected conditions ~25 years in the future. If regionally appropriate, diversification in the form of planting multiple species at each site also appears poised to result in more reliable and resilient reforestation outcomes.